

Overview
Senior Investigator

Research Areas (IRP Lab Groups)
Our main goal is to unravel the nuclear events that drive activation and transformation of B lymphocytes, from changes in transcription, epigenetics and chromatin architecture, to recombination and hypermutation of antibody genes. To achieve these goals, our team combines a wide range of cutting-edge technologies, including genome editing, nanoscopy, in situ Hi-C, cryo-EM, and bioinformatic tools.
B lymphocytes in higher organisms develop in the bone marrow, where they recombine and assemble their cell surface antibody receptor genes. Upon migration to peripheral organs, antibody receptors may bind foreign proteins that belong to pathogens. This recognition leads to a second round of DNA recombination that changes the antibody class, from IgM to IgG, IgE, or IgA. We call this process class switch recombination.
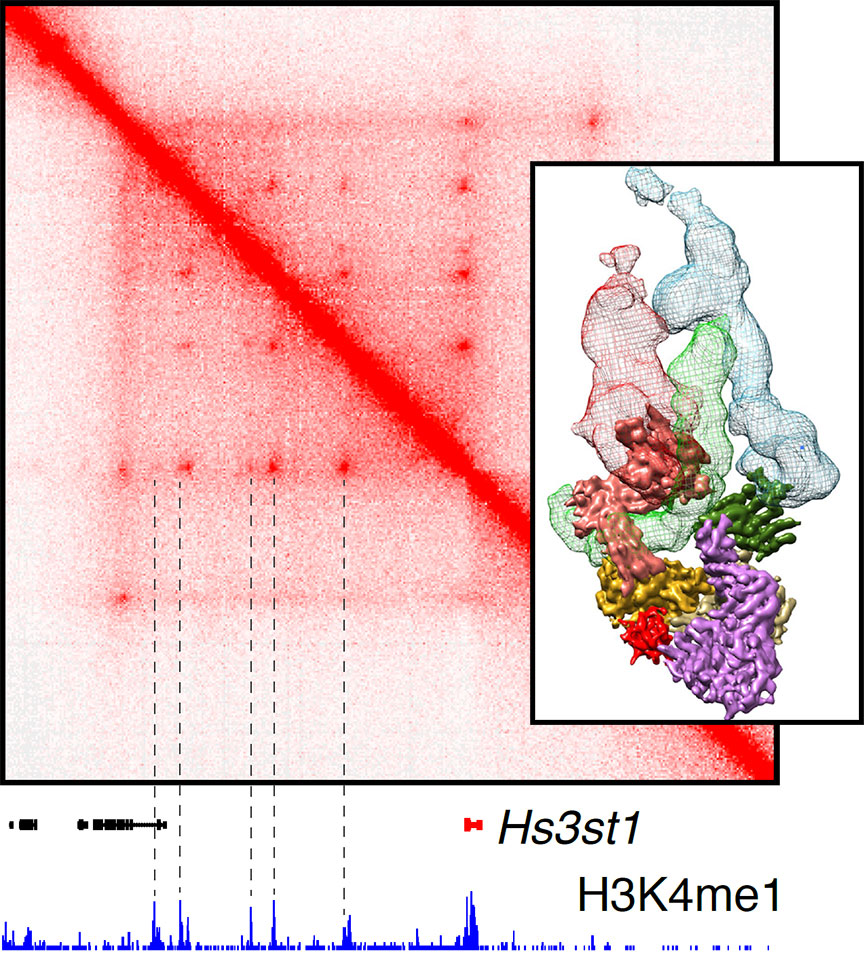
In addition, the antibody genes are hypermutated during the immune response, a phenomenon which coupled to selection improves the capacity of B cells to recognize pathogens.
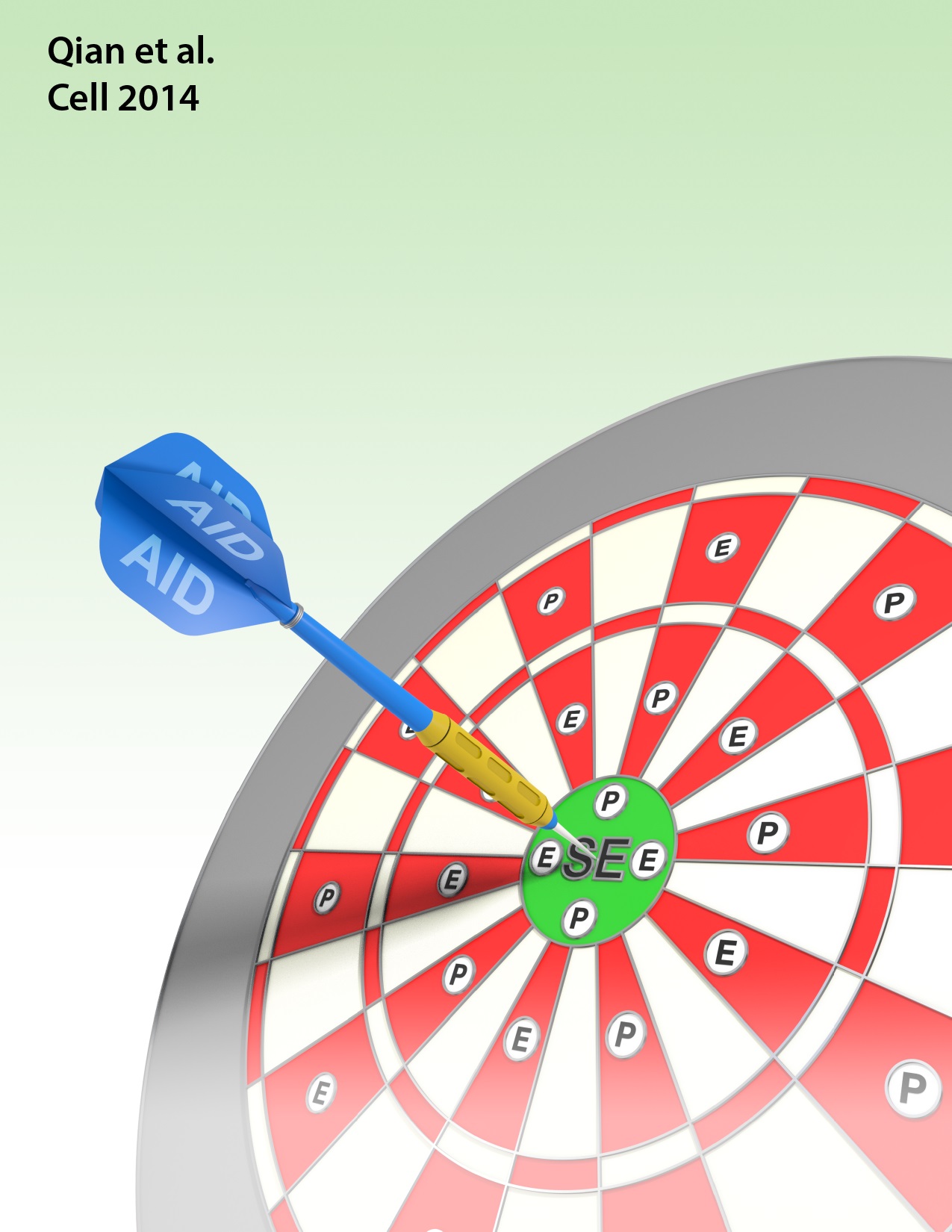
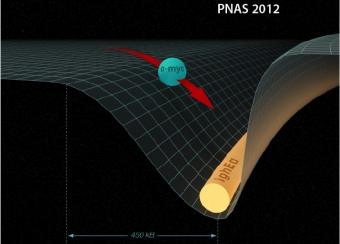
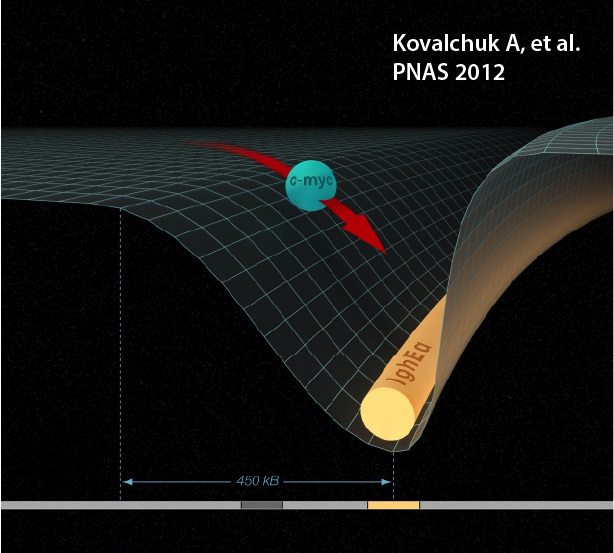
Aside from antibody receptor genes, recombination and hypermutation can promiscuously target other loci in the B cell genome. This can lead to deregulation of transcription and B cell tumor development. Our laboratory has developed several assays to visualize and study such promiscuous activity at susceptible sites.
Our current interests involve:
- Understanding the impact of nuclear architecture on B cell gene expression and transformation.
- Dissecting mechanistic details of chromosomal translocations.
- The production of new mouse models to generate human antibodies of clinical interest.
The Regulome Project
The NIH Regulome Project was established in 2010 as an ongoing collaboration of intramural and invited extramural research groups.
Its primary goal is to understand transcriptional regulation in mammalian systems using cutting-edge technologies, including Hi-C and derivatives, single-molecule microscopy, real-time transcriptional assays, cryo-EM, genomics, biological modeling, CRISPR-Cas9 genetic screens, neural networks, and biophysics.
Core Research Facilities
Labs at the NIAMS are supported by the following state-of-the-art facilities and services:
Staff
Image & Media Gallery


















Scientific Publications
Selected Recent Publications
News & Highlights

